
Die Familie Phrynichidae

Euphrynicus bacillifer Weibchen

Damon annulatipes Männchen
Bestimmung und Anatomie
Die Familie Phrynichidae beinhaltet Arten mittlerer Größe (Phrynichodamon scullyi (20 mm), Arten aus der Gattung Damon Ostafrikas (24-30 mm), Trichodamon (20-28 mm)) sowie sehr groß werdende Arten (Arten aus der Gattung Damon Westafrikas (30-35 mm), manche Phrynichus-Arten (bis zu 35 mm) und Euphrynichus-Arten (bis zu 36 mm).
Morphologisch ist die Familie Phrynichidae als Apulvillata von der Gattung Paracharon, und den Familien Charinidae und Charontidae durch das Fehlen von Pulvilli (siehe Abb. 1, A) an den Tarsi der Laufbeine und das Vorhandensein von Cheliceren mit drei Dentikeln (siehe Abb. 2) in der inneren Dentikelreihe des basalen Chelicerenglieds zu unterscheiden (Weygoldt, 2000).

Abbildung 1: Ventrale Ansichten von Laufbein-Tarsi ohne und mit Pulvilli. A) Tarsus ohne Pulvillus von Euphrynichus bacillifer (Phrynichidae), B) Tarsus mit Pulvillus von Charon grayi (Charontidae). Der rote Pfeil markiert die Stelle, wo sich, wenn es sich nicht um das Laufbein eines Vetreters der Apulvillata handelt, ein Pulvillus befindet. Maßstabsbalken 2 mm.
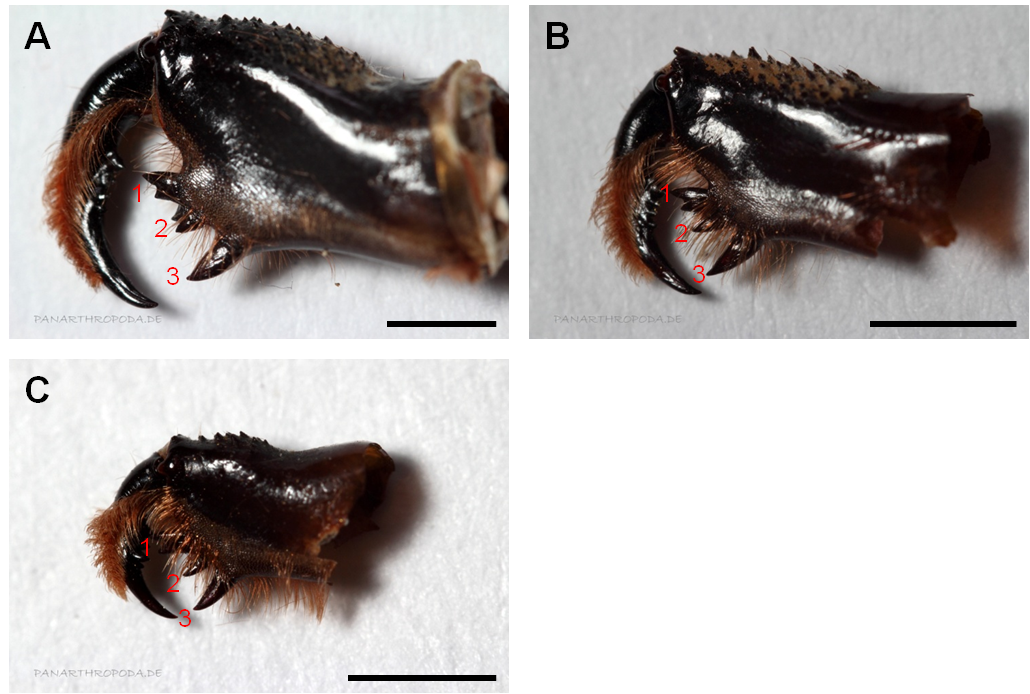
Abbildung 2: Cheliceren von Vetretern dreier Gattungen der Phrynichidae. A) Phrynichus orientalis, B) Euphrynichus bacillifer, C) Damon diadema. Die roten Zahlen markieren die einzelnen Dentikel der inneren Dentikelreihe der Cheliceren. Dentikel 1 ist zweispitzig (bicuspidat), wobei die obere Spitze bei allen Vetretern der Familie Phrynichidae (außer bei Xerophrynus) größer ist als die untere (Weygoldt, 2000). Das in A) gezeigte Dentikel 2 ist nicht zweispitzig, dieser Eindruck erfolgt durch ein weiteres hinter der Dentikelreihe liegendes Dentikel. Zur Aufnahme der Dentikelreihen wurde die Beborstung (teilweise) entfernt. Maßstabsbalken entsprechen 2 mm.
Von den Phrynidae, die diese Merkmale ebenfalls aufweisen, sind die Phrynichidae dadurch zu unterscheiden, dass die beiden mittleren prosomalen Sternite letzterer aus breiten Platten bestehen, die den Großteil der intercoxalen ventralen Fläche bedecken (siehe Abb. 3, A-C). Bei den Phrynidae sind diese Sternite zu kleinen Tuberkel-artigen Strukturen reduziert (siehe Abb. 3, D) (Weygoldt, 2000). Weibliche Genitalien der Phrynichidae besitzen, im Gegensatz zu den Phrynidae, keine klauenartigen Sklerite (Ausnahme bildet hier D. uncinatus) (Weygoldt, 2000).
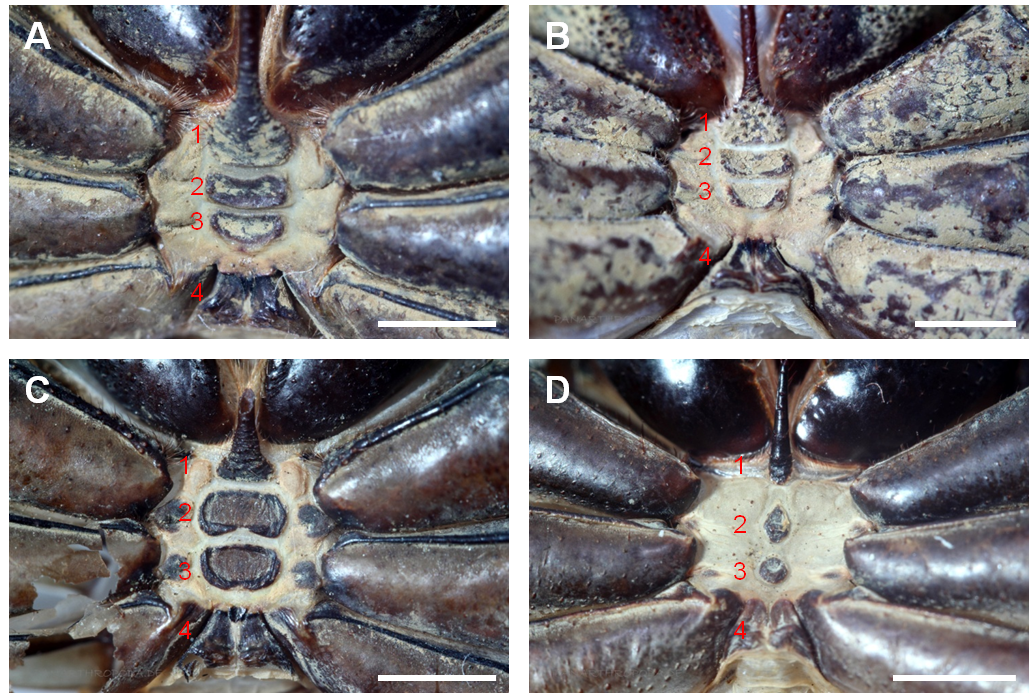
Abbildung 3: Prosomale Sternite dreier Vetreter der Phrynichidae und eines Vertreters der Phrynidae. A) Phrynichus orientalis (Phrynichidae), B) Euphrynichus bacillifer (Phrynichidae), C) Damon diadema (Phrynichidae), D) Phrynus goesi (Phrynidae). Die roten Zahlen markieren die prosomalen Sternite. Bei den Phrynichidae sind die beiden mittleren prospmalen Sternite (2, 3) stark verbreitert und bedecken einen großen Teil der intercoxalen ventralen Fläche (A-C). Bei den Phrynidae (und den restlichen Familien der Geißelspinnen) sind die beiden mittleren prosomalen Sternite zu kleinen Tuberkel-artigen Strukturen reduziert (D). Maßstabsbalken entsprechen 2 mm.
Ein besonderes Merkmal der Familie Phrynichidae (Xerophrynus ausgeschlossen) ist die Änderung des Musters der Bedornung und der Menge an Dornen der Pedipalpen während des postembryonalen Wachstums, welche zur Bildung einer „Fanghand“ führt (Weygoldt, 2000).
Die ersten freilebenden Instars weisen noch die drei primären dorsalen Dornen der Pedipalpen-Tibia auf, welche alle die gleiche Länge aufweisen und für die meisten Vertreter der Neoamblypygi (Charontidae, Phrynidae, Phrynichidae) in diesen Entwicklungsstadien typisch sind (Weygoldt, 2000). Bei Vertretern aus der Familie Phrynichidae wird diese Anordnung im Laufe der Entwicklung und dem Wachstum der Tiere verändert. Die ersten beiden dorsalen Dornen (von distal aus gezählt) der Pedipalpen-Tibia nähern sich einander und verlagern sich an das distale Ende der Pedipalpen-Tibia. Der dritte dorsale Dorn wird oft reduziert oder verschwindet in manchen Fällen ganz. Bei vielen Arten werden die meisten Dornen der Pediplapen-Tibia und -Femur reduziert oder vollständig zurückgebildet (Weygoldt, 2000).
Zusammen mit den ventralen Dornen I u. II der Pedipalpen-Tibia und des Tarsus wird durch diesen Prozess eine für die Phrynichidae sehr charakteristische „Fanghand“ gebildet, die sich vom „Fangkorb“ der Vertreter anderer Familien stark unterscheidet (Weygoldt, 2000).
Beutetiere werden bei den Phrynichidae also durch das Ergreifen mit den „Fanghänden“ gefangen, und nicht wie bei anderen Geißelspinnen durch das Klappmesser-artige Zuschließen der Tibia und des Femur, die durch ihre Bedornung einen „Fangkorb“ bilden.
Viele Arten der Phrynichidae können leicht bezüglich ihres Geschlechtes unterschieden werden. Die Weibchen solcher Arten besitzen an der posterioren Grenze des Genitaloperkulums eine dichte Reihe rötlicher Härchen (siehe Abb. 4) (Weygoldt, 2000).
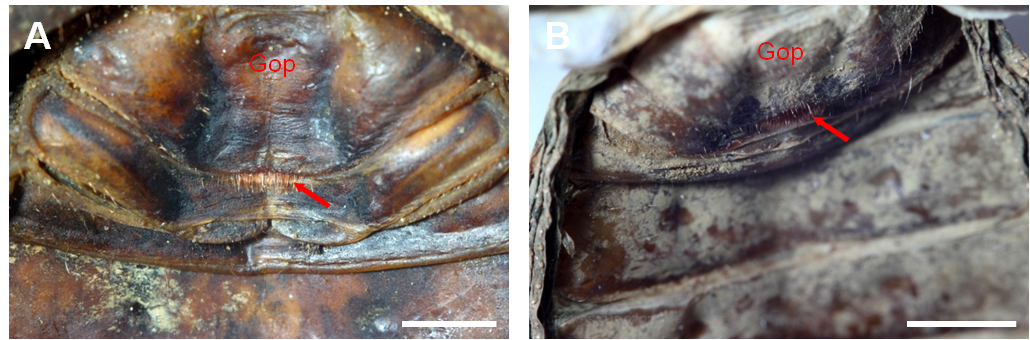
Abbildung 4: Ventrale Ansicht des anterioren Abschnitts des Opisthosomas bei weiblichen Vertretern der Phrynichidae. A) Phrynichus orientalis, B) Euphrynichus bacillifer. Die roten Pfeile markieren die roten Härchen an der posterioren Grenze des Genitaloperkulums (Gop), woran bei vielen Arten aus der Familie Phrynichidae die Weibchen äußerlich von den Männchen unterscheiden werden können. Maßstabsbalken 2 mm.
Verbreitung
Die Familie Phrynichidae ist in Afrika und Asien vertreten, lediglich die Gattung Trichodamon findet sich in Südamerika. Die Gattungen Phrynichodamon, Damon, Musicodamon und Euphrynichus (E. bacillifer ist auch auf Madagaskar vorzufinden) kommen nur auf dem afrikanischen Kontinent vor. Die Gattung Phrynichus kommt in der Palaeotropis vor. Diese Gattung ist in Afrika, den Seychellen, Madagaskar, der arabischen Halbinsel, Indien und Sri Lanka und dem äußersten Osten, wie Kambodscha, Thailand, Malaysia und Indonesien vorzufinden (Weygoldt, 2000).
Es wird angenommen, dass ein Großteil der adaptiven Radiation der Phrynichidae vor dem Auseinanderdriften von Gondwana stattgefunden hat, und dass die heutige geographische Verbreitung der Familie Phrynidae und der Gattung Trichodamon in Südamerika und der Phrynichidae in Afrika und Asien auf die Trennung von Südamerika und Afrika zurückzuführen ist. Somit wäre Trichodamon ein Relikt einer früheren weiteren Verbreitung der Phrynichidae in Südamerika. Möglich wäre jedoch auch, dass die Gattung Trichodamon durch Treibholz nach Südamerika gelangte (Weygoldt, 2000). Die Gattung Damon hat in Afrika eine extensive adaptive Radiation durchgemacht (Prendini et al., 2005; Weygoldt, 2000). Die ebenfalls afrikanischen Gattungen Xerophrynus (deren Einordnung in die Phrynichidae unsicher ist), Phrynichodamon und Musicodamon sind monotypisch (Weygoldt, 2000).
Innere Systematik
Die Familie Phrynichidae besteht aus den Gattungen Xerophrynus, Phrynichodamon, Damon, Musicodamon, Trichodamon, Phrynichus und Euphrynichus. Die Gattung Xerophrynus bildet die Schwestergruppe zu den restlichen Gruppierungen innerhalb der Phrynichidae oder wird als Schwestergruppe neben die Gruppe der Phrynichidae gestellt (Weygoldt, 2000). Traditionell wird die Familie Phrynichidae in zwei Unterfamilien unterteilt (siehe Abb. 5): i) Damoninae, welche Phrynichodamon, Damon und Musicodamon gruppiert und ii) Phrynichinae, in welche die Gattungen Trichodamon, Phrynichus und Euphrynichus gruppiert werden (Weygoldt, 2000). Diese Einteilung ist jedoch unsicher, wobei die Unterfamilie Damoninae ein Paraphylum bilden könnte (siehe Abb. 6), da die phylogenetischen Beziehungen von Musicodamon und Phrynichodamon zu den restlichen Gruppen relativ unsicher ist (Weygoldt, 2000; Weygoldt, 2002). Durch die relativ komplexen Spermatophoren von Musicodamon, die denen der Vertreter der Gattung Trichodamon ähneln, wurde angenommen, dass Musicodamon unter Auschluss der verbleibenden Damoninae, zusammen mit den Phrynichinae eine monophyletische Gruppierung bildet (Weygoldt, 2000).
Eine alternative Erklärung ist, dass komplexe Spermatophoren innerhalb der Phrynichidae eine Synapomorphie darstellen, die die Gattung Damon, Musicodamon und die Phrynichinae, unter Ausschluss von Phrynichodamon, zu einem Monophylum zusammenfasst (siehe Abb. 6). Wenig komplexe Spermatophoren innerhalb der Gattung Damon seien demnach im Laufe der Evolution durch Vereinfachung entstanden, wobei dieser Trend bei D. uncinatus (dessen Spermatophoren nicht bekannt sind, der Bau des weiblichen Genitaltrakts jedoch auf solche zurückschließen lassen könnte) wahrscheinlich nicht stattgefunden hat. D. uncinatus bildet somit wahrscheinlich die Schwesterart zu den restlichen Arten der Gattung Damon (Weygoldt, 2002). Somit müsste in diesem Fall die Unterfamilie Damoninae aufgegeben werden, da Phrynichodamon mit seinen wenig komplexen Spermatophoren und weiblichen Genitalien die Schwestergruppe zu den Phrynichidae mit komplexen oder sekundär vereinfachten Spermatophoren bilden würde (Weygoldt, 2002).
Neuere Erkenntnisse, die genetische Analysen mit einbeziehen, erlauben jedoch womöglich eine Gruppierung der Phrynichidae (mit Ausnahme von Xerophrynus) in Damoninae und Euphrynichinae, wobei die relativ unkomplexen Spermatophoren und weiblichen Genitalträkte von Phrynichodamon ebenfalls auf eine sekundäre Vereinfachung eines komplexeren Bauplans, der dem Vorläufer von Phrynichodamon und Musicodamon gemein war, zurückzuführen ist (siehe Abb. 5) (Prendini et al., 2005).
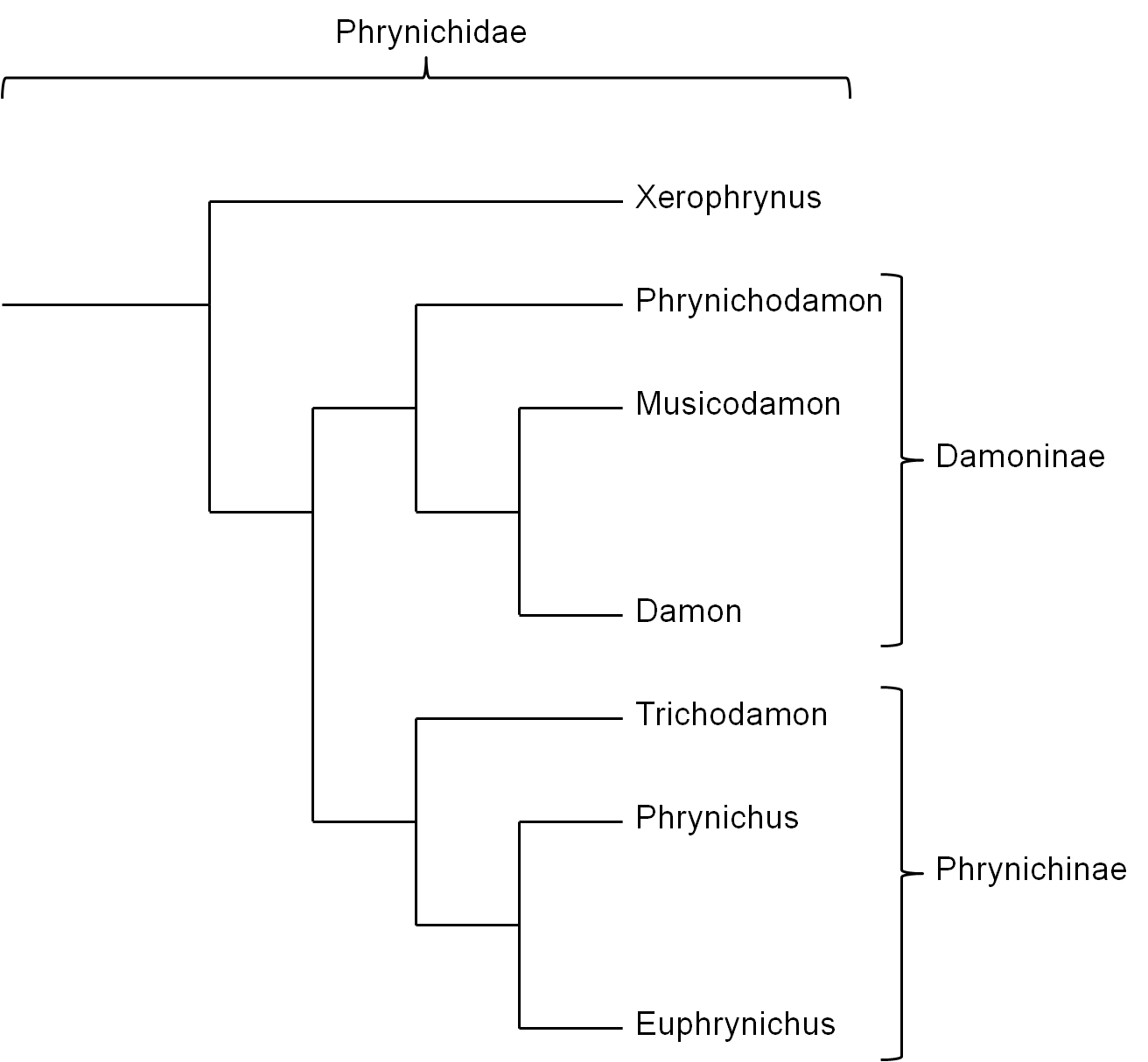
Abbildung 5: Verwandtschaftliche Beziehungen der verschiedenen Gattungen innerhalb der Phrynichidae mit klassischer Einteilung in die Unterfamilien Damoninae und Phrynichinae. Xerophrynus bildet die Schwestergruppe zu allen anderen Vertretern der Familie Phrynichidae (Weygoldt, 2000). Grundlage der Einordnung von Phrynichodamon in die Unterfamilie Damoninae dieses Stammbaums ist die Annahme, dass die relativ unkomplexen Spermatophoren und Genitalien von Phrynichodamon, wie auch bei manchen Vertretern der Gattung Damon, auf eine sekundäre Vereinfachung zurückzuführen sind (Weygoldt, 2002). Komplexe Spermatophoren und weibliche Genitalien sind nach dieser Ansicht also ein symplesiomorphes Merkmal der gesamten Familie Phrynichidae (Prendini et al., 2005; Weygoldt, 2002).
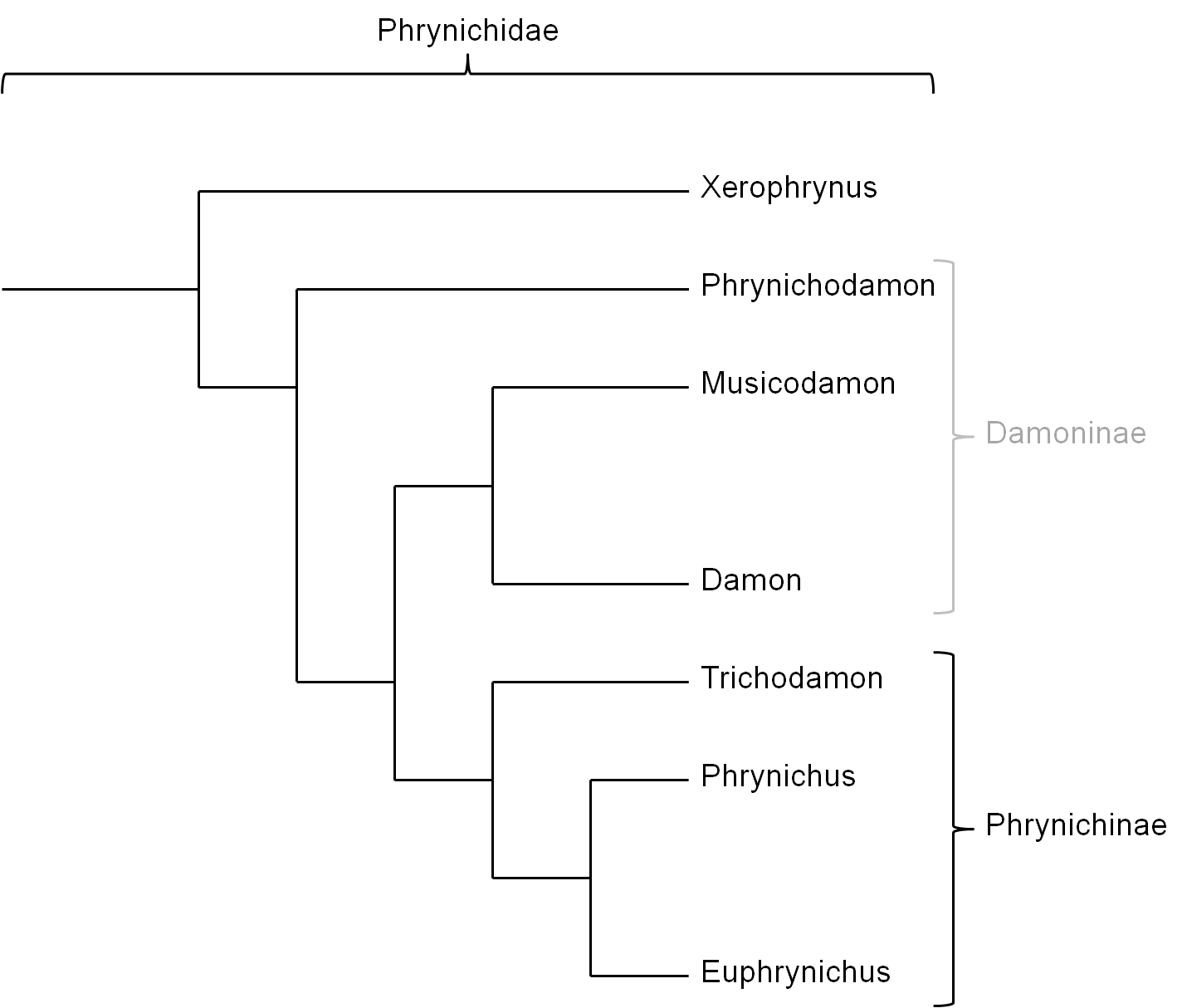
Abbildung 6: Verwandtschaftliche Beziehungen der verschiedenen Gattungen innerhalb der Phrynichidae mit der Unterfamilie Damoninae als paraphyletische Gruppierung. Grundlage für den Ausschluss der Gattung Phrynichodamon aus der somit paraphyletisch werdenden Gruppierung Damoninae ist die Annahme, dass die unkomplexen Spermatophoren und Genitalien von Phrynichodamon keine abgeleiteten sondern plesiomorphe Merkmale sind. Somit sind komplexe Spermatophoren und weibliche Genitalien synapomorphe Merkmale von Musicodamon, Damon (bei diesen oft sekundär vereinfacht) und der Phrynichinae innerhalb der Phrynichidae (Weygoldt, 2002). Die graue Darstellung von „Damoninae“ repräsentiert die Paraphylie dieser Gruppierung unter Annahme der abgebildeten phylogentischen Verhältnisse.
Literatur
Prendini, L. (2005). Systematics of the group of African whip spiders (Chelicerata: Amblypygi): Evidence from behaviour, morphology and DNA. Organisms Diversity & Evolution, 5, 203-236.
Weygoldt, P. (2000). Whips Spiders (Chelicerata: Amblypygi). Their Biology, Morphology and Systematics. Apollo Books, Stenstrup, DK.
Weygoldt, P. (2002). Fighting, Courtship, and Spermatophore Morphology of the Whip Spider Musicodamon atlanteus Fage, 1939 (Phrynichidae) (Chelicerata, Amblypygi). Zoologischer Anzeiger - A Journal of Comparative Zoology, 241, 245-254.
F. Schramm, verfasst 2011